

Сравнение анатомических структур птиц и динозавров
Сравнительная анатомия птиц и тероподов помогла связать их друг с другом. Она также выявила некоторые способы изменения этих характеристик, поскольку динозавры стали более близки к птицам, а птицы оказались более современными. Например, в области таза лобковая кость, первоначально направленная вперед, позже смещается вертикально или назад. В передних лапах относительные пропорции костей оставались довольно постоянными у ранних птиц, но запястье изменилось. У некоторых видов кость в запястье приняла форму полумесяца. В результате ее форма способствовала возможности взлета. Широкая бумерангообразная вилочка у первых оперенных динозавров стала тоньше и образовала более глубокую дугу, когда эта особенность стала необходима для полета.
Недостатки теории Герхарда Хайльмана
Из-за того факта, что у других рептилий имелись ключицы, Хайльман предположил, что тероподы, в частности триасовые псевдозухии, по какой-то причине потеряли их. Для него эта потеря означала, что птицы не могли развиться от этого вида динозавров. Потому он был убежден, как позже выяснилось, ошибочно, что функция ключиц, потерянная во время эволюции, не может быть восстановлена. Птицы, утверждал он, должны были развиться из более архаичной рептильной группы, у которой были ключицы.

Как ранее Сили, Хайльман пришел к выводу, что сходство между птицами и динозаврами должно просто отражать тот факт, что обе группы были двуногими. Его выводы повлияли на палеонтологические исследования и в течение длительного времени были приоритетными, хотя новая появившаяся информация и опровергла некоторые факты. Но до сих пор некоторые ученые придерживаются именно ее. Два отдельных исследования показали, что тероподы действительно имели ключицы. В 1924 году был опубликован анатомический рисунок необычного теропода с попугайным оперением. Он был назван овираптором, и его существование опровергало теорию Хайльмана.

Резюме эволюции морфологии зубов
| Пан троглодит | Sahelanthropus tchadensis | Оррорин тугенский | Ардипитека | Австралопитек афарский | Paranthropus robustus | Paranthropus boisei | Homo habilis | человек прямоходящий | Homo ergaster | Homo heidelbergensis | Homo neanderthalensis | Homo sapiens | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Стоматологическая формула | 2.1.2.32.1.2.3 | 2.1.2.32.1.2.3 | 2.1.2.32.1.2.3 | 2.1.2.32.1.2.3 | 2.1.2.32.1.2.3 | 2.1.2.32.1.2.3 | 2.1.2.32.1.2.3 | 2.1.2.32.1.2.3 | 2.1.2.32.1.2.3 | 2.1.2.32.1.2.3 | 2.1.2.32.1.2.3 | 2.1.2.32.1.2.3 | 2.1.2.32.1.2.3 |
| Образец острия Y-5 | Подарок | Подарок | Подарок | Подарок | Подарок | Подарок | Подарок | Подарок | Подарок | Подарок | Подарок | Подарок | Подарок |
| Расположение куспида | Периферийный | Центральная | Центральная | Центральная | Центральная | Центральная | Центральная | Центральная | Центральная | Центральная | Центральная | Центральная | Центральная |
| Толщина эмали | Тонкий | Средний | Толстый | Средний | Толстый | Очень толстый | Очень толстый | Очень толстый | Очень толстый | Очень толстый | Очень толстый | Очень толстый | Очень толстый |
| Прогнатизм лица | Очень сильный | Очень сильный | Сильный | Сильный | Сильный | Умеренный | Умеренный | Умеренный | Незначительный | Незначительный | Незначительный | Незначительный | Незначительный |
| Форма неба на нижней и верхней челюсти | U-образный | U-образный | U-образный | U-образный | U-образный | U-образный | U-образный | U-образный | U-образный | U-образный | U-образный | U-образный | Параболический |
| Подбородок | Отсутствующий | Отсутствующий | Отсутствующий | Отсутствующий | Отсутствующий | Отсутствующий | Отсутствующий | Отсутствующий | Отсутствующий | Отсутствующий | Отсутствующий | Отсутствующий | Подарок |
| Размер клыков | Очень большой | Большой | Умеренный | Незначительный | Незначительный | Незначительный | Незначительный | Незначительный | Незначительный | Незначительный | Незначительный | Незначительный | Незначительный |
Особенности метода кладистики
Традиционные методы группировки организмов по принципу сходства и различия могут исключить вид из группы только потому, что он имеет черты, которые не были найдены у других членов. В противоположность этому, группы кладистики были основаны исключительно на определенных группах общих черт, которые особенно информативны. Этот метод начинается с дарвиновской заповеди о том, что эволюция продолжается, когда появляется новая наследственная черта в некоторых организмах, которая и передается генетически его потомкам. Предписание указывает, что две группы животных, разделяющие такие новые черты, более тесно связаны друг с другом, чем те, что разделяют только оригинальные, но не производные, переданные по наследству. Определяя общие производные черты, практики-кладистики могут определять отношения среди изучаемых организмов. Результаты таких анализов, которые обычно исследуются, могут быть представлены в виде кладограммы. Древовидная диаграмма, изображающая порядок, в котором появлялись новые характеристики и новые существа, отражают тот порядок, в котором происходила эволюция.
Фон
Сегодня у человека 32 постоянных зуба с зубная формула из 2.1.2.32.1.2.3. Это разбивается на две пары резцов, одну пару клыков, две пары премоляров и три пары коренных зубов на каждой челюсти. У современных людей резцы обычно лопатчатые с одним корнем, в то время как клыки также однокорневые, но с одним бугорком и конической формой. Премоляры двустворчатые, а моляры — множественные. Верхние моляры имеют три корня, а нижние моляры — два корня.
Общие закономерности морфологической эволюции зубов на протяжении всей эволюции человека включают уменьшение лицевого прогнатизм, наличие бугорка Y5, формирование параболического неба и потеря диастема.
Человеческие зубы состоят из дентина и покрыты эмаль в открытых областях. Сама эмаль состоит из гидроксиапатит, а фосфат кальция кристалл. Различные типы зубов человека выполняют разные функции. Резцы используются для разрезания пищи, клыки — для разрыва пищи, а премоляры и коренные зубы — для измельчения и измельчения пищи.
Эволюция зубов у четвероногих позвоночных
Эволюция зубов у сауропсид
Потеря зубов у птиц
Изучение генома различных видов птиц позволяет датировать потерю зубов предками современных птиц 116 миллионами лет назад .
Эволюция зубов у млекопитающих
Регионализация зубов
Первые synapsids были изоморфный зубочелюстной, то есть , чтобы сказать , что все зубы челюсти имели такую же форму, как показано ископаемых пеликозавры ( не- терапсиды synapsids ) как таковые из Dimetrodon . Мы уже можем наблюдать начало регионализации у первых терапсид, в которых мы можем различать резцы , клыки и пост-клыки. Также есть особый зуб, расположенный перед клыком, который называется преканином. Таким образом, возникновение гетеродонтии, то есть дифференциации нескольких типов зубов, имело место у предков терапидов. У млекопитающих продолжается регионализация, и коренные зубы и премоляры отличаются от постклыков. Впоследствии эта регионализация усиливается, и часто развивается дифференциация зубов одного и того же зубного типа, но нового зубного типа не появляется. Однако мы несколько раз наблюдаем потерю одного или нескольких типов зубов, как, например, у монотремов , лишенных резцов и клыков.
Резцы выполняют функцию захвата и стрижки пищи, клыки разрывают пищу, тем самым подготавливая их к растиранию постклыками ( клыковыми зубами ).
Эволюция зубной формулы
Филогенетическое дерево из цинодонты млекопитающих , показывающих эволюцию зубных формул в течение мезозоя . Обозначения узлов (ромбы): 1- Цинодонтия ; 2-млекопитающие; 3-Crown Mammalia ; 4- Монотремы ; 5-эутрихонодонтиды; 6-Trechnotherians; 7-сфалакотероиды; 8-Crown Therians .
Существует общая тенденция к уменьшению количества зубов у млекопитающих. Чаще всего теряются щечные зубы (коренные и премоляры), но часто уменьшается и количество резцов, а клыки выпадают несколько раз самостоятельно. Однако это сокращение не соответствует тому же давлению отбора, которое оказывает на всех млекопитающих, и не было продемонстрировано никакого глобального механизма этого сокращения. Это снижение в каждом случае является адаптацией к диете. Механизмы развития, лежащие в основе потери зубов, не изучены.
Эволюция у людей все еще идет быстрыми темпами, что обычно объясняется снижением давления отбора из-за улучшения доступа к стоматологической помощи, повсеместного приготовления пищи и глубоких изменений в рационе. Кажется, что простое математическое правило характеризует тенденцию (морфогенетический градиент), наблюдаемую у гоминидов с точки зрения размера зубов.
ТОП — Новости мира, инновации
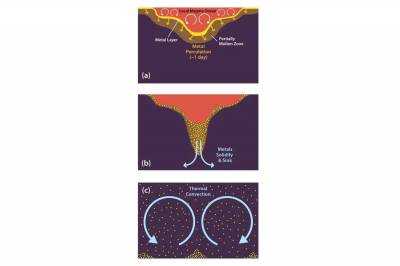 |
| Разработана новая модель происхождения драгоценных металлов в мантии Земли |
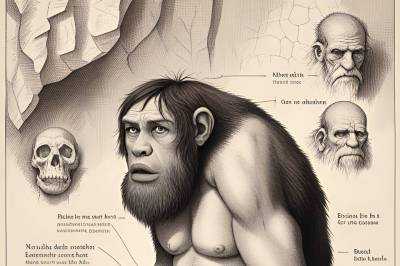 |
| Communications Biology: Гены неандертальцев несут боль современным людям |
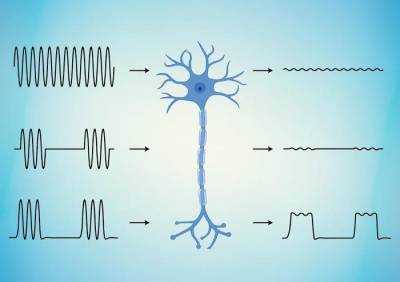 |
| Nature Materials: Ученые создали материал для стимуляции нервной ткани |
 |
| Neuron: Найден новый механизм развития синдрома хрупкой Х-хромосомы |
| Как выглядит работающий рекламный баннер |
 |
| На архипелаге Флорида-Кис открыли улиток ярко-лимонного цвета |
 |
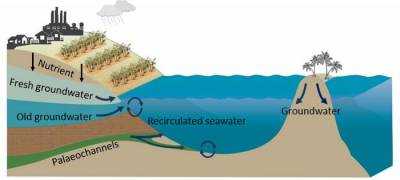
| Датировки человеческих следов в Уайт-Сэндс были точными |
 |
| Ученые предлагают новый подход к сохранению Большого Барьерного рифа |
 |
| Дети, не испытавшие дефицита родительской любви, становятся добрыми взрослыми |
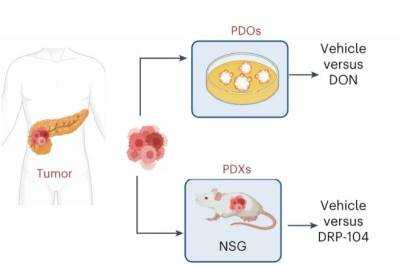 |
| Новый препарат лишает питания раковые клетки поджелудочной железы |
 |
| Current Biology: футбольные вратари обладают уникальным восприятием мира |
 |
| Упасть не беда, беда не подняться: шмели нашли средство против азиатских шершней |
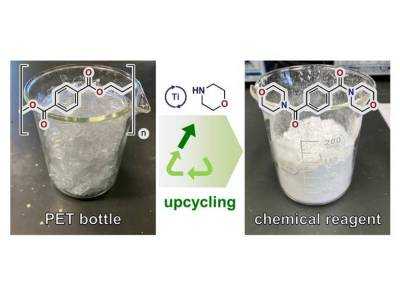 |
| Разработан безотходный и масштабируемый процесс для переработки полиэфиров |
 |
| А не вырубить ли нам все дубы на планете? |
 |
| Создан браслет, отслеживающий перепады настроения при биполярном расстройстве |
Значение истории млекопитающих
Кайнозойская история млекопитающих далеко не такая длинная, как история мезозойских рептилий, но для нас она гораздо важнее. Она прослежена на основе гораздо большего количества ископаемых и представляет более разнообразные виды животных. География и природные условия кайнозоя гораздо ближе к современным, чем мезозойские. Таким образом, нам более знакома обстановка, в которой развивались события. Наконец, млекопитающие принадлежат к той же группе, что и мы сами, и потому представляют для нас большой интерес. В развитии млекопитающих мы более ясно видим возникновение особенностей строения и функций, которые существуют и в нашем теле.
В этой главе мы не останавливались на структурных и функциональных особенностях организмов; мы даже не упомянули приматов, особую группу, к которой принадлежим и мы сами, ограничившись указанием ее места в общей системе на рисунке 60. Приматы для нас настолько важны, что они заслуживают особого рассмотрения. После того как мы рассмотрим природные условия позднего кайнозоя и климатическую обстат новку, создавшую эти условия, мы обратимся в главе восемнадцатой к приматам и посмотрим, к чему они пришли в результате своего развития.
Первые крокодилы триасового периода
До появления первых настоящих крокодилов распространение получили фитозавры — отряд архозавров, очень похожих на крокодилов, за исключением того, что их ноздри располагались на макушке, а не на конце морды. По их названию можно предположить, что фитозавры были травоядными, но это не так. Фитозавры обитали в пресноводных водоемах и охотились на рыбу и другие водные организмы. Среди наиболее известных фитозавров можно упомянуть Rutiodon и Mystriosuchus.
 Фитозавр Rutiodon — не крокодил, но очень похож
Фитозавр Rutiodon — не крокодил, но очень похож
Стоит обратить внимание, что фитозавры, за исключением расположения ноздрей, были больше похожи на крокодилов, чем сами крокодилы того времени. Это еще один пример конвергентной эволюции
Ранние крокодиломорфы обитали на суше и передвигались на задних лапах, некоторые из них были травоядными (предположительно потому, что не могли конкурировать с динозаврами). Эрпетозух и Досвеллия — основные кандидаты на звание родоначальника крокодилов, хотя точных доказательств этого все еще нет. Другой вероятный кандидат — раннетриасовый Xilousuchus, архозавр, который обладал некоторыми чертами характерными для крокодилов.










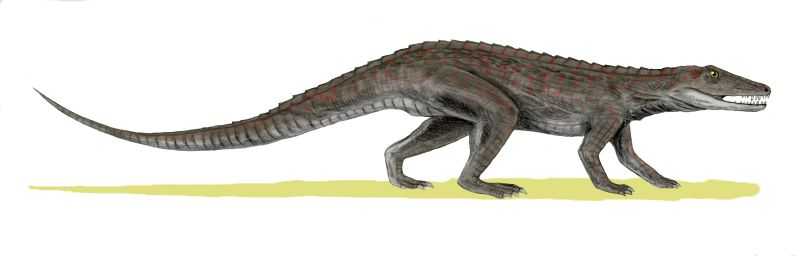 Эрпетозух
Эрпетозух
Как бы то ни было, важно понимать, насколько запутанны и обрывочны палеонтологические свидетельства среднего и позднего триаса. Та часть Пангеи, из которой впоследствии образовалась Южная Америка, кишела динозавроподобными крокодилами, крокодилоподобными динозаврами и ранними птерозаврами, которые были похожи как на крокодилов, так и на динозавров
Только в начале юрского периода динозавры начали развиваться по собственному пути и постепенно стали доминировать в наземных экосистемах. Если бы вы вернулись в прошлое 220 миллионов лет назад и вас бы там съели, то вы вряд ли смогли бы понять кто вас съел: крокодил или динозавр.
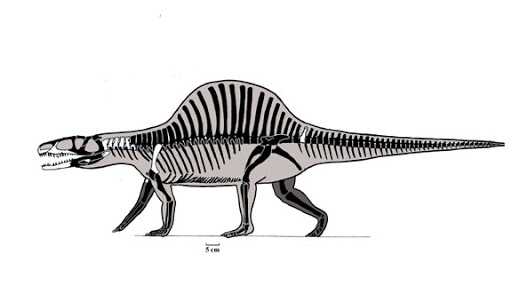 Скелет силозуха
Скелет силозуха
Рекомендации
- ^ Скотт, Г. Р .; Тернер, К. Г. (1997). Антропология зубов современного человека: морфология зубов и ее вариации в недавних популяциях человека. Кембридж и Нью-Йорк: Издательство Кембриджского университета.
- ^
- ^ Скотт, Г. Р .; Ирландский, Дж. Д. (2013). Антропологические взгляды на морфологию зубов: генетика, эволюция, вариации. Кембридж: Издательство Кембриджского университета.
- ^
- ^ Хенке, Винфрид (2007). Хенке, Винфрид; Хардт, Торольф; Таттерсолл, Ян (ред.). Справочник по палеоантропологии: Филогения гоминид. Springer. С. 1527–1529. ISBN 978-3-540-32474-4.
- ^
- ^ Emes, Y .; Айбар, Б .; Ялчин, С. (2011). «Об эволюции человеческих челюстей и зубов: обзор». Бюллетень Международной ассоциации палеодонтологов. 5 (1): 37–47.
- Рак, Ю. (1983). Лицо австралопитека. Нью-Йорк: Academic Press.
- ^ «Результаты оспаривают традиционные представления об эволюции рациона человека». Естественный отбор Newswise.
- ^
- ^ Релетфорд, Дж. (1994). Человеческий вид (2-е изд.). Торонто: Издательская компания Mayfield.
- Schoetensack, O.1908. Der Unterkiefer des Homo heidelbergensis aus den Sanden von Mauer bei Heidelberg. Лейпциг: Вильгельм Энгельманн
Мезозойские млекопитающие
С нашей «позднекайнозойской» точки зрения, современные млекопитающие находятся почти в конце эволюционной линии. Рептилии пермского и триасового периодов, которые походили на млекопитающих, находились на другом конце эволюционной линии. Необходимо помнить, что путь развития организмов от наиболее примитивных млекопитающих до человека не был прямым и это развитие происходило не всегда с одинаковой скоростью. В течение почти 250 миллионов лет мезозоя эволюция млекопитающих не представляла ничего интересного. Можно сказать, что они выжидали своего часа. В то же самое время настоящие рептилии распространились очень широко и приспособились к различным условиям среды в мягком климате. Низменный рельеф и мягкий климат мезозоя способствовали развитию рептилий, но не представляли исключительно благоприятных условий для млекопитающих. Вновь появившиеся млекопитающие должны были выжидать, пока не уменьшились численно и не вымерли динозавры и пока в кайнозое не возникли новые типы природной среды, лишь после чего они в свою очередь смогли сильно распространиться и занять господствующее положение. Все это ясно говорит нам, что эволюционное развитие меняет свое направление в соответствии с меняющимися возможностями, которые предоставляет окружающая среда. Пока происходит движение плит коры и перемещение континентов в более высокие или более низкие широты, а иногда и соединение континентов, будет происходить и изменение условий жизни, создающее новые возможности для эволюции живых организмов.
Вполне вероятно, что предком настоящих млекопитающих мог быть Cynognathus (фото 42) или один из родственных ему видов. А может быть, эта честь принадлежала другой рептилии, еще не обнаруженной в ископаемом состоянии. Это не имеет большого значения, потому что млекопитающие и рептилии, подобные млекопитающим, очень тесно связаны между собой. Какой бы ни была рептилия — родоначальник млекопитающих, — произошедшие от нее млекопитающие имели очень небольшие размеры. Большинство ископаемых млекопитающих, обнаруженных в триасовых, юрских и меловых слоях, были размером с крысу или мышь, а самые крупные не больше кошки. До сих пор не найдено ни одного целого черепа, не говоря уже о скелете, но обнаружено множество мелких челюстей с дифференцированными, как у млекопитающих, зубами и разрозненные кости. Эти кости больше напоминают кости сумчатых или однопроходных, чем плацентарных млекопитающих. В меловых слоях в западной Канаде найдены остатки, принадлежащие, без сомнения, опоссуму.
Небольшие размеры мезозойских млекопитающих указывают на то, что они не могли конкурировать с динозаврами и другими рептилиями, несмотря на имеющийся у них сравнительно большой мозг. Можно предполагать, что млекопитающие занимали подчиненное положение, старались держаться в стороне и, вероятно, питались насекомыми и семенами растений. Так как мы не располагаем целым скелетом, мы не можем представить себе, как они выглядели, но ископаемые черепа позволяют воссоздать внешний вид головы этих млекопитающих (рис. 54).
 Рис. 54. Голова позднемезозойского млекопитающего. Остальные части тела реконструировать пока не удается, так как найден только череп
Рис. 54. Голова позднемезозойского млекопитающего. Остальные части тела реконструировать пока не удается, так как найден только череп
Эволюционные тенденции
Размер тела
Размер тела важен из-за его корреляции с метаболизмом, диетой, историей жизни, географическим диапазоном и скоростью исчезновения. Модальная масса тела динозавров составляет от 1 до 10 тонн в мезозое и во всех основных континентальных регионах. Наблюдалась тенденция к увеличению размеров тела во многих кладах динозавров, включая Thyreophora, Ornithopoda, Pachycephalosauria, Ceratopsia, Sauropomorpha и базальные тероподы. Заметное уменьшение размера тела также произошло в некоторых линиях, но носит более спорадический характер. Самый известный пример — уменьшение размера тела до появления первых птиц; Археоптерикс весил менее 10 кг, а более поздние птицы Confuciusornis и Sinornis были размером от скворца до голубя. Это произошло для облегчения полета.
Мобильность
Древний динозавр был двуногим. Эволюция четвероногой осанки происходила четыре раза, среди предков Euornithopoda, Thyreophora, Ceratopsia и Sauropodomorpha. Во всех четырех случаях это было связано с увеличением размера тела, и во всех четырех случаях тенденция является однонаправленной, без разворота.
Динозавры демонстрируют картину уменьшения и потери пальцев на боковой стороне кисти (пальцы III, IV и V). Примитивная функция руки динозавра — хватать частично противоположным большим пальцем, а не удерживать вес. Уменьшение цифр — одна из отличительных черт тиранозавридов, имеющих только два функциональных пальца на очень коротких передних конечностях.
Влияние источников пищи
Древний динозавр был плотоядным животным. Травоядность среди динозавров возникала трижды, у истоков клад орнитисхиев, зауроподоморфов и теризинозаврид. Отдельные теризинозавриды растительноядны или всеядны. Травоядность среди орнитисхий и зауроподоморфов никогда не была обращена вспять.
Потенциальная совместная эволюция растений и травоядных динозавров была предметом обширных предположений. Появление прозауропод в конце триаса было предположительно связано либо с исчезновением, либо с диверсификацией типов флоры того времени. Появление цератопсид и игуанодонтов и гадрозаврид орнитопод в меловом периоде было предварительно связано с радиацией покрытосеменных. К сожалению, до сих пор нет достоверных данных о диетических предпочтениях травоядных динозавров, кроме данных по технике жевания и гастролитов.
биогеографии
фаун динозавров, которые были относительно однородными по своему характеру, когда Пангея начал распадаться, стал заметно дифференцироваться к концу мелового периода. Биогеография основана на разделении древних видов путем создания географического барьера. Интерпретация ограничена отсутствием окаменелостей восточной части Северной Америки, Мадагаскара, Индии, Антарктиды и Австралии. Не было получено однозначных доказательств биогеографического воздействия на виды динозавров, но некоторые авторы обозначили центры происхождения многих групп динозавров, множественные маршруты расселения и интервалы географической изоляции.
Динозавры, которые были приведены в качестве свидетельства биогеографии, включают тероподов абелизаврид из Южной Америки и, возможно, из других мест Гондваны.
Отношения между динозаврами демонстрируют многочисленные свидетельства расселения из одного региона земного шара в другой. Тероподы-тетанураны широко путешествовали по западной части Северной Америки, Азии, Южной Америке, Африке и Антарктиде. Пахицефалозавры и цератопсы демонстрируют явное свидетельство множественных событий двунаправленной дисперсии в Берингии.
Причина появления перьев у птиц
По мере эволюции птиц и их удаления от динозавров, названных тероподами, множество их особенностей изменялись и совершенствовались. Если присмотреться внимательнее, становится совершенно очевидно, что набор этих характерных признаков долго развивался и служил для выполнения определенной функции. Например, появление оперения у маленького теропода было связано с необходимостью изоляции определенных участков кожи и сначала больше напоминали волосы. На первых птичьих перьях, возможно, присутствовали различные цветовые узоры. Есть версия, что они были предназначены для камуфляжа, распознавания своего вида и других функций.
Примечания и ссылки
- Урсула Лензеле, Пьер Браветти, Эммануэль Гербрант, Филипп Жанвье, « Les dents », La Recherche , n o 407,апрель 2007 г., стр. 75
- Роберт В. Мередит, Гоцзе Чжан, М. Томас П. Гилберт, Эрих Д. Джарвис , Марк С. Спрингер, « Доказательства однократной потери минерализованных зубов у птиц общего предка », Science , vol. 346, п о 6215,декабрь 2014
- Жан Пивето , Трактат по палеонтологии. Происхождение млекопитающих и фундаментальные аспекты их эволюции , Массон,1958 г., стр. 86.
-
(in) ZX Луо , П. Чен , Г. Ли и г-н Чен , Новое эвтриконодонтное млекопитающее и эволюционное развитие ранних млекопитающих , Nature , vol. 446, г.
2007 г., стр. 288-293 ( DOI ) - Brace, CL (1967), Окружающая среда, форма и размер зубов в плейстоцене . J. Dent. Res. 46, 809–816
- Бермудес де Кастро, Дж. М. и Николас, М. Е. (1995) Уменьшение размера передних зубов у гоминидов: доказательства Атапуэрка . Am. J. Phys. Антрополь. 96, 335–356
Ornithischia
Ornithischia, как следует из названия, была придумана для птичьего тазового пояса, хотя они не являются предками птиц.
Орнитисхиальный череп и зубной ряд очень рано изменились за счет травоядной диеты. Лесотозавр отделился рано, но череп Лесотозавра уже демонстрирует такие приспособления с широкими пропорциями, менее гибкой верхней челюстью и более подвижным соединением для нижней челюсти.



![Глава пятнадцатая. млекопитающие [1978 флинт р.ф. - история земли]](https://sttk38.ru/wp-content/uploads/f/5/b/f5b07f6d28fdd8f1aaf74848492857b8.jpeg)






Heterodontosauridae, как было показано, является самой базовой группой в пределах орнитиший. Гетеродонтозавриды очень маленькие (длина тела Абриктозавры все имеют короткий верхний клык и более длинный нижний клык. Передние конечности в известных окаменелостях относительно длинные.
Основные клады были созданы еще в раннюю юрскую эпоху..Орнитисхийцы делятся на бронированных тиреофоранов и небронированных орнитопод и маргоцефалов.
тиреофоранов
Поверхностный доспех (щитки ) самая яркая особенность тиреофоранов. Scutellosaurus имеет это, но в остальном мало отличается от лесотозавра. У него длинный хвост и комбинированная поза двуногих и четвероногих, которая отличает его от всех более поздних тиреофоров, включая Стегозаврия и Анкилозаврия. Эти две клады, хотя и совершенно разные по внешнему виду, имеют много общих черт черепа и скелета.
Стегозавров легко узнать по выступающему ряду пластин над позвоночником и длинных шипов на хвосте. ауры, но не стегозавр, также имеют по спине на каждом плече. Эти шипы и пластины произошли от более ранних поверхностных щитков. Хуаянгозавр — самый старый и самый примитивный известный стегозавр.
Анкилозавров легко узнать по их обширному доспеху. Череп сильно окостенел. В начале своей эволюции анкилозавры разделились на Nodosauridae и Ankylosauridae, которые отличались особенностями черепа.
Ornithopoda
Ornithopods делятся на отдельные клады — Hypsilophodontidae и Iguanodontia.
Hypsilophodontids больше напоминают своих предков, чем гетеродонтозавриды. Наиболее отличительные черты — короткая лопатка и предлобковый отросток палочковидной формы. Самый ранний из них — Agilisaurus из средней юры Китая.
Игуанодонты — это разнообразный, но морфологически тесно связанный ряд родов, известных по окаменелостям позднего мела. К значительным изменениям относится эволюция зубных батареек, связочная пясть и положение пальцев рук. Тенонтозавр — самый базальный игуанодонт. Другие включают Iguanodon, Camptosaurus и Muttaburrasaurus.
Marginocephalia
Marginocephalia, названные по названию выступа над задней частью черепа. Они включают пахицефалозавров и цератопсов.
. Пахицефалозавры наиболее известны своими толстыми верхними частями черепа. Самый старый из известных — Stenopelix, относящийся к раннему меловому периоду Европы.
Цератопсы, известные благодаря Протоцератопсу, Трицератопсу и Стиракозавру, иллюстрируют эволюцию черепов с оборками и рогами. Оборки произошли от полочки, общей для всех Marginocephalia. Цератопсы делятся на базальные цератопсы, включая клювых попугаев Psittacosaurus и неоцератопсов.
Разнообразие цератопсов черепов. А) Скелет Протоцератопса. Б) к I) Черепа. B) и C) Пситтакозавр сбоку и сверху. D) и E) Протоцератопс сбоку и сверху. F) и G) Трицератопс сбоку и сверху. H) и I) Стиракозавр сбоку (без нижней челюсти) и сверху.
Эволюция цератопсидных динозавров имеет общие черты с эволюцией некоторых групп млекопитающих, оба были «геологически короткими» событиями, ускоряющими одновременную эволюцию большого размера тела, производных структур питания и «разнообразных роговидных органов».
Последовательность эволюции цератопсов в меловом периоде примерно от Пситтакозавра (121-99 млн лет) до протоцератопса (83 млн лет) до (трицератопс 67 млн лет и стиракозавр 72 млн лет). На виде сбоку череп Пситтакозавра очень мало похож на череп Стиракозавра, но на виде сверху можно увидеть аналогичное пятиугольное расположение.
Новые доказательства
В 1936 году Чарльз Камп из Калифорнийского университета в Беркли нашел останки небольшого теропода раннего юрского периода и хотел поставить точку в истории с ключицами. Но доказательство ошибочности теории Хайльмана было мало кем признано. Недавние исследования нашли ключицы у многих тероподов. Исходя из строения птиц в настоящее время, а также в результате ряда исследований, археоптерикс был назван промежуточным звеном в истории эволюции пернатых. Но оставался вопрос: между какими именно рептилиями? Дальнейшие ископаемые птицы найдены в слоях ниже раннего мела. Большинство из них были обнаружены с начала 1990 годов, главным образом в Китае. Между тем известно более тридцати видов птиц из мелового периода. Кроме того, было обнаружено большое число маленьких динозавров, передвигающихся на двух ногах с характерными для древних птиц чертами. Таким образом, разрыв между динозаврами и птицами благодаря изучению окаменелостей стал значительно меньше.
Начало исследования возникновения птиц
Ученые принялись размышлять об эволюции птиц, начиная с того времени, как Чарльз Дарвин изложил свою теорию эволюции в работе «О происхождении видов». В 1861 году, год спустя после публикации трактата Дарвина, древнее птичье перо было найдено в баварских известняковых отложениях, возраст которых составлял около 150 млн лет. В следующем году был найден скелет животного, у которого были птичьи крылья и перья, очень длинный костлявый хвост и челюсть с зубами. Он был найден в том же регионе. Окаменевший скелет был назван археоптериксом. Он стал первым найденным древнейшим животным с оперением. Скелетная анатомия археоптерикса давала четкие доказательства, что предками птиц были динозавры, но в 1861 году ученые еще не смогли установить эту связь. Тогда и началось изучение эволюции птиц и охота за предками современных видов пернатых.
Общие признаки динозавров и птиц
Исследования Готье, проведенные в последнее время, демонстрируют, что многие функции, которые традиционно считаются принадлежащими только птицам, фактически появились до них, у их предков — тероподов. Многие из этих свойств помогли своим первоначальным обладателям выжить. Эти же черты и некоторые другие были в конечном итоге использованы или преобразованы для полета и жизни на деревьях. Птичьи характеристики тероподов, которые эволюционировали до птиц, не проявлялись сразу, а некоторые присутствовали до того, как появились сами тероподы. Они были еще у динозавров, существовавших ранее. Например, непосредственный предок тероподов был двуногими и передвигался как птица. Он был маленьким и плотоядным. У него были передние лапы, как и у ранних птиц. Кроме того, второй, а не третий палец, как у других рептилий, был длиннее. У предков динозавров голеностопный сустав стал шарнирным, а кости стопы — удлиненными. Многие из изменений в строении ног, как полагают, повлияли на увеличение длины шага и скорость бега. Это свойство в один прекрасный день поможет птичьим тероподам взлететь.

Теория Острома
Наконец, спустя столетия после спорной презентации Хаксли для Геологического общества Лондона, Джон Х. Остром из Йельского университета возродил идею о том, что птицы были связаны с тероподами. Он прямо предположил, что пернатые являются их прямыми потомками. В конце 1960-х годов Остром описал скелетную анатомию хищного теропода дейнониха, который был размером с человеческого подростка и обитал на Земле около 115 миллионов лет назад, в эпоху раннего мела. В дальнейших своих опубликованных работах Остром продолжил определение анатомии птиц и группы особенностей, которые они, включая археоптерикса, делили с дейнонихом и другими тероподами, но не с другими рептилиями.









На основе этих данных он пришел к выводу, что птицы происходят непосредственно от маленьких динозавров-тероподов. После выдвижения теории Остром продолжал собирать свои доказательства происхождения пернатых от тероподов. Он использовал новый метод расшифровки отношений между организмами, применяя его в музеях естественной истории в Нью-Йорке, Париже и других местах. Этот метод получил название кладистики или филогенетической систематики. Поскольку он стал стандартом для сравнительной биологии, его использование строго обосновывало выводы Острома.
